
嘉峪检测网 2025-06-10 16:01
导读:除了对每批次的单克隆抗体进行常规的放行检测之外,对使用临床试验样品生产工艺生产的产品进行进一步的生物化学和生物物理学上的表征,对于全面了解单克隆抗体产品的详细结构 特征,并鉴定关键质量属性是至关重要的。这些详细的表征数据还可以用于支持产品制剂处方的 开发,有助于证明工艺放大、工艺变更或者厂房变更后的产品可比性。不需要对每批次的单克隆抗体
除了对每批次的单克隆抗体进行常规的放行检测之外,对使用临床试验样品生产工艺生产的产品进行进一步的生物化学和生物物理学上的表征,对于全面了解单克隆抗体产品的详细结构 特征,并鉴定关键质量属性是至关重要的。这些详细的表征数据还可以用于支持产品制剂处方的 开发,有助于证明工艺放大、工艺变更或者厂房变更后的产品可比性。不需要对每批次的单克隆抗体都进行表征研究,而且本文中所述的分析方法也并非都需要在临床I 期研究之前进行。
用于表征单克隆抗体产品的分析方法应尽可能完整的描述产品的结构和变化。从表4.9 中 列出的常用的表征单克隆抗体产品的分析方法的清单中可以看出,某些分析方法既可以用于产品 的表征,也可以用于放行测试。比如,肽图分析可以用作常规的鉴定测试,因为它可以确证单克 隆抗体产品的一级结构。同时,肽图还可以提供有关单克隆抗体的更为详细的表征信息,比如抗 体中二硫键的确认,是否存在降解产物,如蛋白质的氧化或者脱酰胺形式,以及各种翻译后的位 置和结构修饰,比如糖基化。另一种常用于放行和表征的分析方法是质谱分析。除了确认正确的 分子量,蛋白序列和翻译后修饰之外,质谱分析还可以用于鉴定单克隆抗体存在其他产品相关或 者不相关的蛋白,并为肽图分析确定的产品结构信息提供正交的确认。
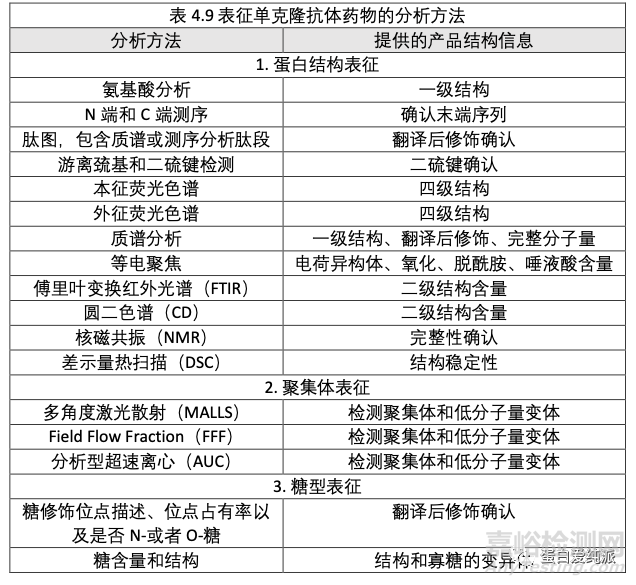
1 蛋白结构的表征
1.1 氨基酸序列分析
氨基酸分析可以鉴定和定量蛋白质样品中存在的大多数的氨基酸,样品通常要经过酸水解 法破坏各个氨基酸之间的肽键。酸水解通常使用6 N 的盐酸在100℃下处理16-24 小时。释放单 个氨基酸后,可以在色谱分离之前或者之后进行衍生化,然后进行在线检测。通过与同时运行的 标准品进行比较来鉴定和定量氨基酸。尽管这是大多数蛋白质分析实验室中常用的常规方法,但 是此方法也有缺点,就是对某些氨基酸分析不准确。含有酰胺键的氨基酸,比如天冬酰胺和谷氨 酰胺,会被转换为相应的酸形式,天冬氨酸和谷氨酸,而无法最终单独测定。一些其他的氨基酸, 比如丝氨酸、苏氨酸、半胱氨酸和甲硫氨酸,会因为苛刻的降解条件而发生部分降解,因此无法 准确定量。一种氨基酸,色氨酸,会被常用的酸水解方法而破坏。
1.2 N 端和C 端序列分析
证明单克隆抗体具有正确的N 端和C 端是确认产品的一级结构以及确定是否存在不同形式 加工的抗体形式的另一种分析方法。C 端的赖氨酸变体和N 端的前导肽序列的不完全切割是单克 隆抗体产品常见的变异体,这种变异体可以通过N 端和C 端序列分析来检测的。这些变异体是可 接受的,但是要进行检测以确保在整个工艺开发过程中,不同规模的或者不同的生产工艺生产的 单克隆抗体产品批次之间的一致性。如果N 端没有被谷氨酸残基环化形成的焦谷氨酸封闭,那么 通常可以使用标准的Edman 降解方法对完整单克隆抗体进行N 末端测序。通常要测定重链和轻 链的N 端5-10 个氨基酸残基,并将所得的序列与单克隆抗体产品产物的预期序列进行比较。测定C 端的氨基酸序列则通常需要将蛋白质降解为小肽,然后对小肽进行测序,这里面即存在重链和 轻链的C 末端。通常作为整个单克隆抗体肽图分析的一部分来进行。
相比于单条链的蛋白质,单克隆抗体产品中含有两条蛋白链使得对序列数据的解释更加困 难。另外,无法使用Edman 降解法对焦谷氨酸测序,因此通过该方法无法检测到含有焦谷氨酸的 重链的N 端序列,但是可以在肽图中确定C 端序列。重链序列的缺失不能证明焦谷氨酸的存在, 但这是最可能的原因。因此,N 端测序通常为定性检测,以确认是否存在预期的序列,而不是对 每个存在的序列进行定量。在一些或者全部重链的N 端含有焦谷氨酸的情况下,可以通过肽图分 析来确定N 端序列。
1.3 肽图分析
如上面所讲,肽图分析可以用来确认单克隆抗体产品的一级结构,但是它也可以鉴定因降 解,比如脱酰胺或者氧化,而导致的单个氨基酸变化。肽图分析还可以用于确定单克隆抗体产品 中二硫键的位置和糖基化位点。单克隆抗体产品的肽图分析可以视为蛋白质的指纹图谱,提供了 对该蛋白质的全面的理解。
当用于表征研究时,肽图分析应确认至少包含了90%的蛋白质序列,单克隆抗体的肽图分 析可能使用不止一种酶来消化蛋白质。与序列相关,单克隆抗体的某些部分可能无法被酶消化, 从而导致肽段太大而无法被鉴定。另外的部分也有可能被彻底消化成了单个氨基酸,这些氨基酸 在分离肽段的色谱中无法保留。翻译后修饰的存在还可能是抗体的某些部分对酶消化产生抗性, 因此需要在消化和肽图分析之前将蛋白质进行去糖基化处理。
1.4 游离的巯基和二硫键
在正常的条件下,IgG1 单克隆抗体含有四个链间二硫键,其中两个将轻链与重链连接,两 个将两条重链彼此连接。其他IgG 的链间二硫键的数目略有不同,但是所有的IgG 均通过二硫键连 接在一起。在可变区和恒定区也会发现额外的链间二硫键,它们有助于稳定单克隆抗体的三维结 构。尽管单克隆抗体上所有的半胱氨酸残基都应以二硫键配对,但是一个或多个二硫键还是可能 被还原,从而导致单克隆抗体产品中游离的巯基含量较低。这些游离的巯基可与另一个抗体分子 中的游离巯基反应,导致二聚化和多聚化,从而影响单克隆抗体此产品的稳定性。
单克隆抗体产品中的半胱氨酸残基是否配对成二硫键可以通过在还原和非还原条件下产品 的肽图分析来确定。将单克隆抗体去糖基化,去除蛋白质主链上的碳水化合物后,其中的一份样 品用4- 乙烯基吡啶烷基化, 胰蛋白酶或者其他合适的酶消化, 经过HPLC 分离进行质谱分析 (LC/MS)检测。另一份的样品先用二硫苏糖醇(DTT)还原,用4-乙烯基吡啶烷基化,用相同的 酶消化并通过LC / MS 分析。通过比较所得的两个肽图,可以鉴定出含有通过二硫键配对的半胱氨 酸残基的肽段。对于单克隆抗体产品,含有许多的二硫键,通过还原前收集分离的肽段,可以更 容易的确定在二硫键中配对的半胱氨酸残基。对还原和非还原两种情况获得肽图分析,可以清楚 的确定哪个两个半胱氨酸配对成了特定的二硫键。
正确折叠的单克隆抗体产品不应该存在游离的巯基。可以通过使用Elman 试剂(DTNB)处 理单克隆抗体产品来确定产品中的游离巯基的存在程度。Elman 试剂与单克隆抗体中的游离巯基 反应,生成对硝基苯酚,该物质在412nm 处有非常强的吸收。在进行DTNB 处理之前,需要使用 温和的变性剂处理来破坏单克隆抗体产品的三维结构,以将所有的游离的巯基暴露于Elman 试剂。反应结束后,测定在412nm 处的吸光度值,扣除412nm 处的背景吸收,然后除以13600(对硝基 苯酚的消光系数),以此来定量存在的游离巯基。Elman 试剂可以用许多其他的反应试剂来代替, 这些反应试剂都是特异的结合游离巯基,然后转换为比色或者荧光测定,即使存在过量的未结合 试剂情况下仍可定量游离巯基含量。
1.5 内源性和激发性荧光色谱
内源性荧光光谱(IFS)可以提供有关单克隆抗体产品的三级结构的定量信息。在这个方法 中,蛋白质被近紫外到蓝光波长的激光激发,可诱导单克隆抗体中的色氨酸残基发出特征性的荧 光,这反映了每个色氨酸残基周围的微环境。单克隆抗体产品的固有荧光对蛋白质的三级结构 非常敏感,因为变性、降解或者聚集导致的三级结构变化都会导致固有荧光的最大波长或者强度 发生变化。因为三级结构与固有荧光之间没有绝对的关系,因此不可能从荧光信息中得到抗体结 构的信息。但是,该方法在评估强制条件下蛋白质三级结构的变化中非常有用,并且经常用于分 析抗体的强制降解样品,作为抗体产品稳定性研究的一部分工作。该方法可以用于制剂研究和对 生物类似药的评估。
也可以通过使用与某些氨基酸(比如色氨酸)特异性结合的荧光探针来定量激发性荧光。但是探针的结合可能会破坏单克隆抗体的结构,因此还是倾向于使用内源性荧光测量作为研究单 克隆抗体三级/四级结构的方法。
1.6 核磁共振
核磁共振波谱(NMR)通过检测原子核之间的相互作用来测定相互作用的结构信息。NMR 通过分析每个质子旋转引起的磁化强度来提供蛋白质原子分辨率级别的结构信息。NMR 分析高度 依赖于技术和软件,因此需要昂贵的设备和大量的计算机分析才能对数据进行反卷积,然后确定 哪些氨基酸侧链对于抗体大分子的相互作用有影响。如果可以使用C13 或者N 标记抗体,则有助 于抗体的研究。蛋白质的标记通常可以通过在富含这些同位素化合物的培养基中培养微生物来 实现。尽管NMR 不常用研究单克隆抗体的结构,但是可以用于研究抗体与目标分子之间的相互作 用。比较结合前后的NMR 光谱有助于确定哪些氨基酸侧链是接近结合于结合区域的。
NMR 最大的优点是可以用于分析溶液中的蛋白质的结构,尽管溶液中蛋白质浓度要求要足 够的高(0.05-1.0 mM)才能获得有用的结果。这是一种非破坏性的方法,意味着样品可以进行 二次分析。NMR 的分析结果可以快速获得,尽管只是部分获得,但是非常有用的。目前NMR 还 无法分析分子量大于40kDa 的蛋白质,因此通常用于分析小分子量的蛋白质。
1.7 质谱
质谱分析(MS)是通过多种方式在蛋白质上产生电荷,然后通过磁场和/或电场加速带电 的蛋白质来分离抗体。蛋白质的加速度正比于其质荷比,然后在检测器中被俘获。蛋白质的电 离可以通过稀有气体的原子轰击(快速原子轰击,FAB),激光电离(基质辅助激光解离,MALDI) 或者电喷雾电离(ESI)来实现。还有其他可以使抗体蛋白质带电荷的方法,但是上述这些是最常 见的。带电离子通过分析仪加速,通常是飞行时间(TOF)或者四级杆类型。飞行时间(TOF)质 谱仪是指可以在电场中加速带电蛋白质,从而使每个具有相同电荷的分析具有相同的能量。分子 的飞行速度仅仅取决于蛋白质的质荷比。每种分析类型都有不同的适用范围,适用于某些不同的 电离方法。比如,TOF 分析仪可以适用于ESI 或者MALDI,而四极杆分析仪则不适用于MALDI 电离 样品。检测器通常为光电倍增器或者电子倍增器。质谱仪的类型通常使用电离器和分析器类型来 描述,比如MALDI-TOF 质谱。检测器记录捕获的带电粒子,并根据加速模式来计算质荷比(m/z), 通过合适的计算机软件,就可以将其转换为原始分子的质量。MS 可以和诸如高效液相色谱(HPLC) 之类的分离方法组合使用,先将样品的组分进行分离,然后确定每种组分的分子量。之前,这种 方法费时又费力,限制了其实用性。但是,现代质谱仪以及配合其解释结果而开发的软件系统 使得该方法更易于使用,目前已经得到广泛的应用。氢氘交换(HDX)功能标记和片段分离技术 可以与MS 结合使用用来鉴定蛋白质表面残基。
质谱分析可以提供有关抗体分子量的准确信息,但是必须要注意选择合适的方法来使蛋白 质离子化,并选择加速离子化物质的条件。能量过高的电离可能会导致单克隆抗体分解为重 链和轻链,甚至有可能片段化抗体。质谱仪的输出结果包含一系列的峰,代表了所检测到的质量, 但是峰的高度与单独的蛋白质的含量并不成比例,因此该方法无法提供定量检测的结果。质谱图 中的峰高与离子数量成正比,但是离子数量与蛋白质的离子化程度、离子化形式的稳定性以及通 过质谱仪加速的程度有关。为了能够定量各种蛋白质,需要将MS 分析与输出结果与蛋白质含量 成正比的方法,比如反相HPLC 结合使用。这种分离方法需要开发工作,以便使其适用于MS 检测 以及大多数的HPLC 方法所用的UV 检测。通常用于反相HPLC 的三氟乙酸不适用于MS 样品,因此 需要开发其他的缓冲液系统。
除了确定抗体的完整分子量之外,还可以通过使用激发能使抗体片段化来获得其序列的数 据。使用两个直接连接的质谱仪(MS/MS),以足够的能量来轰击蛋白质,从而使抗体产生多个 蛋白质片段。这些片段被加速,然后单独的片段再使用额外的能量轰击产生二级片段,在第二个MS 中被加速和检测。使用专门的软件,可以通过检查所检测到的片段并发现哪些片段与已知单独 氨基酸的分子量片段的区别来确定肽段序列。这些都可以足够快的完成,以便可以在第二次激发 中检测第一次激发中产生的大多数片段。当今MS/MS 设备的高质量和高灵敏度甚至可以提供有关 氧化或者脱酰胺以及一级结构方面的信息。
1.8 等电聚焦
上面描述的等电聚焦分析方法,如果不用于产品的放行检测,则通常用于蛋白质的表征研 究。该技术具有非常高的分辨率,能够分离仅有一个电荷差异的蛋白质至单独的条带中。因此, 该方法非常适合用于测定单克隆抗体产品的电荷异构体,还可以监测产品的脱酰胺、氧化和唾液 酸化程度。尽管该方法可以测定抗体的电荷异质性,但是没有办法来验证异构化的位点,必 须与另一种方法,比如肽图分析,结合使用来鉴定氧化或者脱酰胺的位点。
1.9 圆二色谱
圆二色性(CD)是一种光吸收现象,代表的是左右圆偏振光的差异吸收。圆二色谱(CD) 就是基于分子,比如抗体在远紫外区域(190-250nm)或近紫外区域(230-250nm)影响左右偏振 光的方式的差异。这些差异的产生就是因为抗体,和所有的蛋白一样,在结构上的不对称性。单 克隆抗体,如果没有有序的结构,则不会产生强度信号。有序的结构则可能产生正的或者负的CD 信号。从这些区域的CD 光谱上获得的信息是不同的。远紫外CD 对二级结构更为敏感,而近紫外CD 光谱对三级结构更为敏感。
CD 可以用于测定蛋白质的二级结构。通常可以用于确定折叠或者突变对蛋白质的影响以 及用来研究蛋白质之间相互作用。在远紫外区域,α-螺旋,β-折叠或无规卷曲通常具有良好并且 独特的CD 光谱。单克隆抗体产品在远紫外区域的CD 光谱则反映了蛋白质结构中的每个光谱所代 表的蛋白结构的数量。该方法还可以用于评估抗体微环境的改变如何影响其二级结构。这些微环 境可包括不同的缓冲液pH 或者溶液的渗透压。与远紫外CD 不同,近紫外CD 光谱与特定的蛋白 结构之间没有直接的关联。近紫外CD 光谱区域取决于芳香族氨基酸,比如酪氨酸、苯丙氨酸和色 氨酸、半胱氨酸的吸收和其微环境。CD 一般不常用于放行测试,但是CD 可用于比较不同批次的 单克隆抗体的二级结构,还可以用于制剂处方和制剂工艺的开发。
CD 是一种样品无损的分析技术,可以对样品进行再分析。它要求样品的浓度要稍高,如0.5-1.0mg/ml。CD 可以快速的获得结果,并且可以在各种条件下,比如不同的pH 和温度下进行研究。单克隆抗体产品的分析很大程度上依赖于已知结构的蛋白衍生的参照光谱的经验,因此, 一个成功的CD 分析是取决于所使用的参考数据库的。
1.10 傅立叶变换红外光谱
傅里叶红外光谱(FTIR)也是用于分析蛋白质的二级结构的。红外光谱法测量入射能量与 分子内的电偶极相互作用时在特定能量下吸收的入射红外辐射(0.78-1000um)。FTIR 依赖于红 外光谱学基础,也就是蛋白质二级结构与相关红外波段条带之间的关系。FTIR 仪器与先进的数学分析方法相结合,可以分离重叠的IR 波段。FTIR 提供了有关抗体蛋白高度结构化区域,比如α螺 旋或β折叠的存在和相对数量的信息。结构化区域影响抗体的酰胺键区域的红外光谱的吸收。通过 对已知二级结构含量的蛋白质的研究结构分析,可以将FTIR 数据与各种结构域的含量相关联。FTIR 相对于其他检测二级结构的分析方法一个优势是它对固体制剂的适用性。该方法还可以应用 于冻干或者喷雾干燥的样品来评估多种制剂处方。
1.11 多角度激光散射
多角度激光散射(MALLS)提供了有关单克隆抗体产品聚集体和降解体的数量和性质的信 息。对于聚合物,比如PEG 偶联的抗体,MALLS 也可以很好的估计其真实大小,而SDS-PAGE 或者SEC 是不能看到的。
1.12 差示量热扫描
单克隆抗体也可以通过差示量热扫描方法(DSC)来进行表征研究。与FTIR 类似,DSC 提 供的是有关抗体热稳定性的信息。通过测定影响样品相变所需的能量,DSC 提供了有关不同制剂 中的单克隆抗体的比较信息。折叠成热力学最稳定结构的单克隆抗体要比结构不稳定程度高或者 具有不稳定的三级/四级结构的抗体需要更多的能量来展开。相变温度(Tm),也就是样品中50% 的分子未折叠点的温度是产品三级结构稳定性的函数。单克隆抗体的折叠和解折叠发生在分子中 的多个位置,从而导致会有多个热转变点。由于单克隆抗体产品的Tm 可能会受到产品所用的制 剂的影响,因此DSC 也可以作为筛选和优化单克隆抗体产品制剂处方的重要工具。
1.13 表征蛋白结构的其他分析方法
尽管在临床I 期之间很少使用,但是还是有其他的几种分光光度法可以用于表征单克隆抗 体产品。测定UV 光谱的二阶导数可以提供有关抗体纯度的信息。FTIR 和CD 可以提供有关抗体的 二级结构的信息。每种单克隆抗体的芳香族氨基酸(酪氨酸、色氨酸和苯丙氨酸)的含量可能不 同,从而导致该抗体在近紫外区域(250-350nm)内的特征光谱有所不同。通过测定光谱的二阶 导数,可以确定该光谱是由于单个纯分子还是两个或者多个光谱组合的结果,从而表明是否存在 杂质。
另一种常见的降解途径是天冬酰胺的脱酰胺,可以通过市售的试剂盒来测定48 。该方法确 定异天冬氨酸的含量主要是基于当异天冬氨酸与异麦芽糖基甲基转移酶(PIMT)一起时可以酶促 催化S-腺苷甲硫氨酸(SAM)转化为S-腺苷高半胱氨酸(SAH)。SAH 的含量即等于转化的异天冬 氨酸的含量,然后通过HPLC 进行分离和检测。通过与同时运行的标准曲线来确定SAH 的浓度。该 方法适用于定量天冬酰胺的脱酰胺含量,因为异天冬氨酸主要通过脱酰胺产生。不过要注意的是, 由于溶液的pH 和周围的氨基酸序列可能会影响生成的异天冬氨酸和天冬氨酸的比例。
2 聚集体的表征
单克隆抗体产品原液和制剂中的聚集体是影响产品活性和安全性的关键质量属性。上文讨 论的尺寸排阻色谱法和SDS-PAGE 可用于常规的检测单克隆抗体聚集体。为了更详细的表征单克隆 抗体产品,精细的分析方法,比如多角度激光散射(MALLS)、分析型超速离心(AUC)和非 对称快速超滤(FFF)等,则可以使用。
MALLS 利用的溶液中的蛋白质对已知光散射的行为。以光源为直线,从零角度散射的光量 与溶液中的蛋白质的质量成正比。零角度光因为被光源淹没,无法测量,因此MALLS 仪器测量的 是一定角度(>3)散射的光,并使用此数据来计算溶液中分子的平均分子量。该方法能够在单一 溶液中鉴定出多种不同分子量的分子,因此不需要事先使用SEC 来分离抗体。MALLS 也可以用于 确定从SEC 分离出的峰上分子量,这就可以提供有关分离的高分子量形式的分子是二聚体、三聚 体还是多聚体形式的信息。为了获得有关单克隆抗体聚集体形式的性质和数量的最多信息, 将SEC 与UV 和 MALLS 检测结合使用既可以定量蛋白质的峰,又可以鉴定出每个峰中的分子的分子量 信息。
AUC 是基于蛋白质或者其他聚合物在离心场中通过梯度溶液的迁移率来分析。该迁移率是 所研究抗体的分子半径的函数。尽管AUC 不适合于质量控制,而且由于其精度差而易于验证,但 是在确定单克隆抗体及其相关产品形式的分子量时,AUC 仍是一个金标准,同时也可以用于确定 抗体的结合常数。AUC 数据分析的前提是基于单克隆抗体产品分子半径与其分子量时直接相关的。只需要少量的样品,AUC 即可分析共价和非共价的聚集体。尽管可以确定较大的分子量范围,但 是在高浓度抗体溶液中,AUC 的检测并不是特别理想,因此需要稀释样品,这可能会导致可逆聚 集体的解离。AUC 数据的分析需要专门的软件将沉降速率转换为分子量信息,而对数据的解释则 需要专门的人员培训。
非对称快速超滤也可以同时分析出共价和非共价的聚集体。FFF 分离是通过施加垂直于样 品通过狭窄通道的流场将溶质分层固定在膜上来实现。分子沿着流动路径的运动是所分离的分子 的分子量的函数。由于没有柱分离,因此该方法不能分离非常大的分子,并且难以看到0.15um 范 围内的聚集体和颗粒 73 。与AUC 一样,该方法的精密度较差,难以验证,不适合QC 使用。样品 与膜之间的相互作用现在尚未有很好的理解和控制,可能是导致结果不准确的原因。
3 糖型的表征
单克隆抗体产品表征的一个关键部分是确定分子中的碳水化合物含量,包括单糖的数量和 类型,确定蛋白质上碳水化合物链的定位以及单克隆抗体上存在的各种聚糖的寡糖模式,确定每 个糖链结构中末端存在的唾液酸含量和类型。确定单克隆抗体产品上存在的寡糖是N 糖或者O 糖也非常重要。哺乳动物细胞生产的单克隆抗体通常在每条重链的CH2 区域的Asn297 上存在 一个N-连接的糖链结构,但是,这种糖基化的程度会因为不同的生产细胞系而变化,并且可能取 决于用于生产单克隆抗体产品的培养条件。此外,CHO 细胞还可以在单克隆抗体上引入人抗体上 不存在的糖或者糖链,从而产生非活性或者具有免疫源性的产物。在单克隆抗体产品的其他天 冬酰胺残基上也会发现类似的N-连接糖链。比如, 目前批准的单克隆抗体产品,如西妥昔单抗 (Erbitux)约有15-20%的VH 区域的Asn88 上具有N-连接的糖基化。由于抗体糖基化是生产过 程中批间差异的主要原因,监管机构要求全面表征单克隆抗体产品的糖基化性质和程度。
单克隆抗体产品的完整糖基化表征包括分析分子中每一个潜在糖基化位点的百分比以及确 定不同的寡糖的结构。如上所述,肽图分析可以用于鉴定单克隆抗体产品中的糖基化位点以及位 点的占用比例。利用MS/MS 或者N 端测序分析对肽图分析中释放的肽段进行分析,可以鉴定出产 物中每个糖基化位点的确切位置。每个位点的占有率则可以通过测定含有糖基化位点的肽段在有 或者没有糖链的情况下的相对含量来确定。进行此类分析时,需要注意确保糖链的存在不会对蛋白质的消化降解产生影响。通过对释放的每种肽段进行测序,可以在某种程度上补偿这种情况, 从而可以鉴定出切割位点的差异。
描述每一个糖基化位点的寡糖结构可能会比较复杂,因为每个糖型结构可能包含不同的含 量的唾液酸,可能存在双触角或者三触角形式,也可能被岩藻糖化等。确定每种糖型的结构首先 使用PNGaseF 从单克隆抗体上切割下完整的寡糖,然后通过HPLC 分离不同的寡糖结构,使用一种 或者多种方法进行测定。每个寡糖的结构信息可以通过将样品的洗脱时间与在相同色谱条件下同 时运行的一系列标准寡糖进行比较来确定。或者可以通过质谱法来确定寡糖的结构,类似于单克 隆抗体产物的肽链上的氨基酸测序。将此方法确定的测序数据与先前确定的单糖组成数据相结合, 可以确保序列的准确性。
4 亚可见颗粒的定量
单克隆抗体产品中存在的潜在颗粒的数量和大小对产品的一致性和安全性至关重要,尤其 是随着FDA 越来越关注尺寸小于10um 的颗粒。在这个尺寸范围内,药典上用于测量颗粒的方法 (USP <788>和EP 2.9.19)准确性要稍差。尽管尚无监管机构期望控制此尺寸范围的颗粒物,但是 已经开始要求企业要确定此尺寸范围内的颗粒物的浓度并汇报结果。当前药典方法测定这些小颗 粒时的不准确性,还需要在测定中应用其他的分析方法。新方法可包括图像分析、激光衍射和 偏振强度衍射分析(PID)。图像分析可以捕获悬浮在溶液中的颗粒经过感应区域时的图像。图像 分析软件可以确定颗粒的浓度和大小。激光衍射分析基于与MALLS 相同的原理。入射光的微分散 射是溶液中颗粒大小的函数。偏振强度衍射也是使用光的微分衍射作为颗粒大小的函数关系来分 析,但是入射光在垂直和水平两个方向上偏振,并且在多个波长下测定散射光以增加所获得信息。目前,这些方法都不适合在QC 实验室中实施,但是每种方法都可以提供有关单克隆抗体产品中存 在的亚可见(≤10um)的微粒的大小和浓度的信息。

来源:Internet
关键词: 单克隆抗体